How gradients of chemical signals that shape animal bodies evolve
The question of how the enormous range of animal diversity that surrounds us came to exist has fascinated scientists for centuries. Since the 1930s, Charles Darwin’s theory of evolution by natural selection has been broadly accepted as the process that shapes new species. Many people, myself included, usually imagine natural selection only during an organism’s adult lifetime – say, one finch’s beak might have superior digging power, allowing it to outcompete its fellow finches and pass on this trait to the next generation. However, an entire field of science studies evolution by examining variations in embryonic development between species, an area of work known as evolutionary developmental biology.
Two UChicago researchers in this field, Urs Schmidt-Ott, associate professor of Organismal Biology and Anatomy, and Chun Wai Kwan, a graduate student in the Schmidt-Ott lab, along with their colleagues Edwin L. Ferguson, professor of Molecular Genetics and Cell Biology, and postdoctoral scholar Jackie Gavin-Smyth (a former graduate student in the Ferguson lab), have provided evidence for a novel evolutionary mechanism during development in a study recently published in eLife.
As an embryo develops, chemical signals determine how distinct tissues form over time and space. Bone Morphogenetic Proteins (BMPs), found in organisms from fruit flies to humans, belong to a class of signaling molecules known as morphogens, essential factors in directing the arrangement of tissue types as an embryo grows. By comparing the distribution of BMP signaling during the development of two different fly species, Schmidt-Ott and his team have identified the BMP gradient as a target for evolution.
During development, the fly embryo’s cells spit out BMPs into the extracellular space, where they diffuse around the embryo. Cells lining the back, or dorsal, region of the embryo carry receptors that bind the BMPs, which in turn sets off chemical signals within the cells that drive tissue specification. Generally, the intensity of BMP signaling follows a gradient pattern: most intense at the dorsal region and decreasing in magnitude in cells spread further to the sides. In Drosophila melanogaster, the fruit fly commonly used as a model organism, cells experiencing high BMP signaling early in development become an extraembryonic tissue called amnioserosa, which plays important roles in protecting the embryo and directing the movement of tissue in late developmental stages.
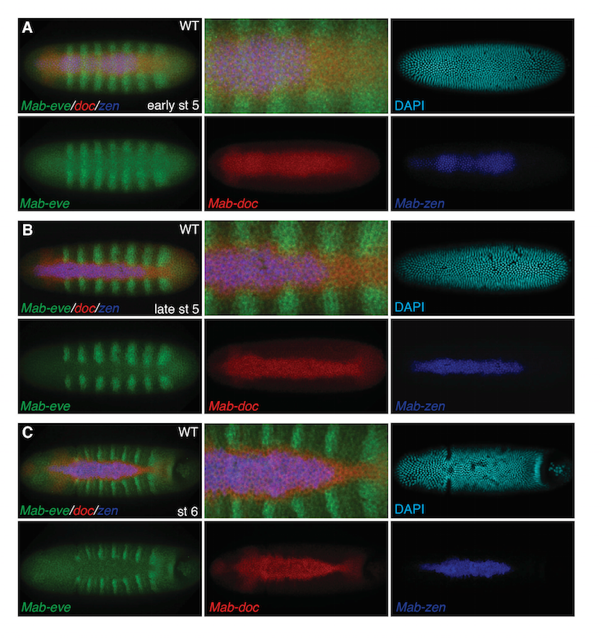
However, in most insects, including many fly species, BMP signaling promotes the formation of two separate tissue types, the serosa and the amnion. These extraembryonic tissues fulfil functions similar to those of the amnioserosa, as well as mediating key elements of embryo immunity. InMegaselia abdita, a fly closely related to Drosophila, the serosa develops from the most dorsal region and the amnion from surrounding tissue. As Schmidt-Ott, Kwan, and their team demonstrate in the new paper, the BMP signaling gradient in Megaselia first becomes narrow and steep, and then broadens as development progresses. By observing and manipulating levels of tissue-specific genetic markers as well as the changing BMP gradient in Megaselia embryos, they found that a positive feedback loop in pre-amnion cells amplifies BMP signaling after the serosa has begun forming, widening the range of BMP signaling and thus determining amnion.
Timing as well as spatial distribution of BMP signaling is key for the formation of distinct tissue types. In Megaselia, early low-level BMP signaling activates genes in nested domains that mark, but do not specify, the two extraembryonic tissue types. Rather, target genes of the early BMP gradient function in a positive feedback loop, first concentrating BMP signaling in the prospective serosa and then– after serosa specification – in the prospective amnion. In Drosophila, with its single extraembryonic tissue, the BMP gradient also becomes narrow and steep in the process of amnioserosa specification, but fails to broaden thereafter.
These results suggest that at some point in its evolution, Drosophila lost the broadening of the BMP gradient. Rather than identifying a gene downstream of the BMP gradient that directly produces a trait that has changed between species, this study indicates that genes responsible for the spatiotemporal dynamics of the BMP gradient during development were targeted in evolution to alter tissue. In Kwan’s words, “we found that by changing the shape of the morphogen gradient, the morphology or tissue type in different species can also change. This is a different way of looking at how evolution can occur.”
The subtlety of the interplay between extracellular signals like BMP and the responding cells, and the implications in determining animal forms demonstrated by this work support a theory of body patterning proposed by Alan Turing, the famous mathematician, in the 1950s. A simple model of signaling in development posits that a field of cells responds to a static signaling gradient. In this model, evolutionary change of tissue types or the organization of the animal’s body plan would be mediated by changing the nature of the responding cells.
As Schmidt-Ott explains it, Turing instead imagined that pattern and form depend primarily on the form of the morphogen gradients. Turing envisioned multiple diffusing signaling molecules interacting with each other in a concentration-dependent manner, and thereby defining their respective gradient shapes. By modeling the diffusion and reactivity of morphogens mathematically, he illustrated how a stable pattern can emerge from random fluctuations. From Turing’s consideration of the importance of a dynamic signaling gradient in animal body plans, the potential for a change in a morphogen gradient to advance evolution naturally follows.
“What was interesting for us here is that it was the morphogens themselves that were driving the evolutionary process,” said Schmidt-Ott. “By changing the parameters by which they are distributed in time and space, you get different animal forms.”
Next, Kwan and Schmidt-Ott plan to examine a group of flies that have two tissue types, likeMegaselia, but have a reversed morphology towards the end of development – the amnion tissue ends up on the ventral side, rather than the dorsal as in Megaselia. They plan to determine whether different signaling gradients play a role in determining the ultimate arrangement of tissues as well as specifying tissue types, and to determine the genetic elements involved. They plan to pursue this goal – once again – in close collaboration with the Ferguson laboratory to combine their work with new experimental with research in Drosophila.
Schmidt-Ott hopes that these results will inspire the consideration of signaling gradients as targets of evolution more broadly.
“We could imagine this being applicable to the neural tube of vertebrates or the eyespots on butterfly wings – the same principle in which morphogens control the evolution of the complexity of a trait through feedback loops,” he said. “As evolutionary targets, feedback loops would change the dynamic of these gradients and that would provide a mechanism for diversification of the trait in question.”
Kwan and Schmidt-Ott would like to acknowledge the contributions of the additional authors on the paper, Jackie Gavin-Smyth, a postdoctoral scientist in the Reinitz lab, and Edwin L. Ferguson, professor of Molecular Genetics and Cell Biology, whose work on body patterning in Drosophila provided insight into the signaling pathway that promotes formation of amnioserosa tissue.